
Synthetic community [SynCom] transfer for the...

Find out more about the different routes to entry and our eligibility criteria
Yet frogs employ a wide range of locomotor modes, with several taxa featuring primary locomotor modes other than jumping. Using a combination of techniques (CT imaging and 3D visualization, morphometrics, phylogenetic mapping), this study aims to determine the link between skeletal anatomy and locomotor style, habitat type and phylogenetic history, shedding new light on how functional demands impact morphology.
Body and limb measurements for 164 taxa from all the recognised anuran families are extracted from digitally segmented CT scans of whole frog skeletons and analysed using various statistical techniques. We find that the expansion of the sacral diapophyses is the most important variable for predicting locomotor mode, which was more closely correlated with frog morphology than either habitat type or phylogenetic relationships. Predictive analyses suggest that skeletal morphology is a useful indicator of jumping but less so for other locomotor modes, suggesting that there is a wide range of anatomical solutions to performing locomotor styles such as swimming, burrowing or walking.
The history of anuran morphometrics is dominated by a focus on the relationship between limb lengths and jumping performance (Rand, 1952; Zug, 1972; Dobrowolska, 1973; Choi et al., 2003; James et al., 2005; James et al., 2007; James & Wilson, 2008; Herrel et al., 2016). In contrast, other types of locomotor modes, including walking, hopping, swimming, burrowing and climbing, have received comparatively less attention (Emerson, 1979; Wells, 2007; Robovska-Havelkova et al., 2014; Vassallo et al., 2021), as have regions of the body other than the pelvis and hindlimb such as the forelimb, hands and feet (Manzano et al., 2008, 2019; Keeffe & Blackburn, 2020, 2022; Abdala et al., 2022).
Despite their relatively conserved body plan (Lires et al., 2016), extant anurans show considerable modifications in their skeletal proportions, reflecting their ability to respond to various mechanical challenges and inhabit diverse environments (Citadini et al., 2018; Gomes et al., 2009; Moen, 2019; Moen et al., 2013; Simons, 2008; Soliz et al., 2017; Vidal-García et al., 2014). What remains unclear is how the interplay between morphology, function, ecology, and phylogeny drove the evolution of these modifications (Buttimer et al., 2020).
The total length of the hindlimb, and how it compares to forelimb and body length, is a good predictor of jumping performance, including jumping distance and take-off speed (Choi et al., 2003; Emerson, 1978; Gomes et al., 2009; James & Wilson, 2008). While both terrestrial and arboreal jumpers have long hindlimbs, arboreal jumpers are said to have similarly elongated forelimbs to meet the biomechanical requirements for both climbing and jumping (Simons, 2008), or to compensate for the potential problem of a displaced centre of gravity (De Oliveira-Lagôa et al., 2019). Similarly, more equally elongated forelimbs and hindlimbs have been shown to be associated with walking (Reynaga et al., 2018).
However, comparatively fewer studies have considered whether individual hindlimb segments are associated with different functions during locomotion (Dobrowolska, 1973; Enriquez-Urzelai et al., 2015; Lires et al., 2016). Additionally, the thickness of the forelimb relative to its length may potentially correlate with fossoriality, whereby proportionally larger humeral crests afford broader attachment sites for forelimb muscles used in digging (Emerson, 1976; Keeffe & Blackburn, 2020). Similarly, hindlimb thickness in aquatic species is associated with large muscles used for underwater propulsion (Gillis & Biewener, 2000). Significantly, however, a lack of detailed comparative investigations of limb ratios may be hampering conclusions about how locomotor function and skeletal proportions covary.
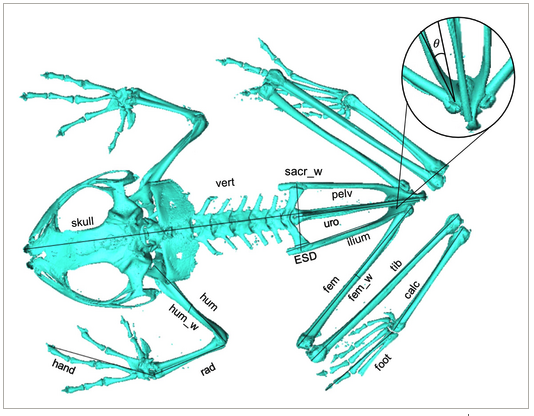
In addition to limb proportions, pelvic morphology has also been linked to variations in locomotor performance and habitat use (Emerson, 1979, 1982; Prikryl et al., 2009; Pugener & Maglia, 2009; Reilly & Jorgensen, 2011; Jorgensen & Reilly, 2013; Soliz et al., 2017; Buttimer et al., 2020). Pelvic specializations include the shape and degree of expansion of the sacral diapophyses (ESD), the presence or absence of dorsal ridges on the ilia and urostyle, and the morphology of the sacral-urostylic joint. These features are widely used to identify three pelvic types, each associated with a specific type of movement, namely: ‘lateral-bending’, ‘fore-aft sliding’ and ‘sagittal-hinge’. Previous studies have proposed that these pelvic types occur in walker-hoppers, swimmers and jumpers, respectively (Emerson, 1979, 1982; Jorgensen & Reilly, 2013; Reilly & Jorgensen, 2011).
More recently, it has been demonstrated that frog families (Manzano & Barg, 2005) and locomotor groups do not fall neatly into these groups (Simons, 2008; Soliz et al., 2017). Whether Emerson's three pelvic types accurately represent species-level variation in pelvic morphology remains uncertain, suggesting that Emerson's concept of pelvic types should be revisited with a broader analysis.
